At my day job we work with Node 12 for our projects, but I wanted to check out NuxtJS 3, which requires version 14 or 16. Once I updated npm, I found out me old projects no longer worked properly. Reinstalling node_modules didn’t solve the problem, and besided, I didn’t want to saddle up my colleagues with version incompatibilities I couldn’t detect myself. So, I looked into finding a solution. And found one!
With n you can switch easily between Node versions. To install:
npm install -g n echo "export N_PREFIX=/home/jeroen/.local/n" >> ~/.bashrc echo "export PATH=$N_PREFIX/bin:$PATH" >> ~/.bashrc
Don’t install it as root, that will make it much more annoying to work with.
Now it is easy to change between versions:
n 16 node -v >> v16.13.1 n 14 node -v >> v14.18.2
But still, you would have to remember which version is used in which project. That’s not fun at all, so that’s what avn was made for. I guess I’m not the only one who is annoyed by such things.
To install avn:
npm install -g avn avn-nvm avn-n avn setup
That last command adds a line to ~/.bash_profile, which is ignored in my setup, so I had to move that line to ~/.bashrc :
echo '[[ -s "$HOME/.avn/bin/avn.sh" ]] && source "$HOME/.avn/bin/avn.sh" # load avn' >> ~/.bashrc
Now add a .node-version file to the root of your project (or in my case the root of all the projects of my boss):
echo "12.22.8" >> ~/projects/.node-version
I also did the same for my own project (with version 16), and now when I switch to any child folder of ~/projects I see this:
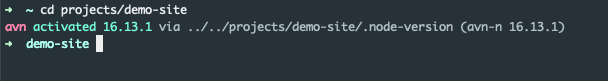
Mission accomplished!
Troubleshoot
While installing this to Macos, I ran into a bit of trouble:
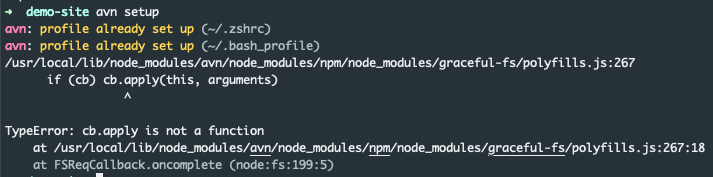
If found this solution for it:
n 10.13.0 # Install node version 10.13.0 by n rm -R ~/.avn nvm exec 10.13.0 npm install -g avn avn-nvm avn-n # Use installed version to install the packages nvm exec 10.13.0 avn setup
This only works with already installed node versions. When I went to a directory with a node version that isn’t installed yet, I got this:
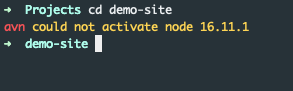
My current solution is to just install that version manually with n:
n 16.11.1
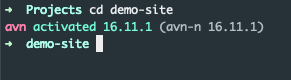
Mission accomplished again!
Recent Comments